
Search results
There is a page named "Plant hormone" on Wikipedia
 Plant hormones (or phytohormones) are signal molecules, produced within plants, that occur in extremely low concentrations. Plant hormones control all...56 KB (7,035 words) - 10:31, 16 September 2024
Plant hormones (or phytohormones) are signal molecules, produced within plants, that occur in extremely low concentrations. Plant hormones control all...56 KB (7,035 words) - 10:31, 16 September 2024 plant hormone. It is the simplest alkene gas and is the first gas known to act as hormone. It acts at trace levels throughout the life of the plant by...33 KB (3,843 words) - 12:48, 6 July 2024
plant hormone. It is the simplest alkene gas and is the first gas known to act as hormone. It acts at trace levels throughout the life of the plant by...33 KB (3,843 words) - 12:48, 6 July 2024 and behavior. Hormones are required for the correct development of animals, plants and fungi. Due to the broad definition of a hormone (as a signaling...40 KB (4,214 words) - 20:12, 18 August 2024
and behavior. Hormones are required for the correct development of animals, plants and fungi. Due to the broad definition of a hormone (as a signaling...40 KB (4,214 words) - 20:12, 18 August 2024 Salicylic acid (redirect from Salicylic acid (plant hormone))precursor to and a metabolite of acetylsalicylic acid (aspirin). It is a plant hormone, and has been listed by the EPA Toxic Substances Control Act (TSCA)...39 KB (3,477 words) - 13:03, 21 October 2024
Salicylic acid (redirect from Salicylic acid (plant hormone))precursor to and a metabolite of acetylsalicylic acid (aspirin). It is a plant hormone, and has been listed by the EPA Toxic Substances Control Act (TSCA)...39 KB (3,477 words) - 13:03, 21 October 2024 Shine Muscat (section Treatment with plant hormone)producing seedless or large fruits using gibberellin, a type of plant growth hormone, was developed in Japan. This method has been adapted to Shine Muscat...12 KB (1,356 words) - 07:22, 14 September 2024
Shine Muscat (section Treatment with plant hormone)producing seedless or large fruits using gibberellin, a type of plant growth hormone, was developed in Japan. This method has been adapted to Shine Muscat...12 KB (1,356 words) - 07:22, 14 September 2024 physiologists study fundamental processes of plants, such as photosynthesis, respiration, plant nutrition, plant hormone functions, tropisms, nastic movements...30 KB (3,553 words) - 20:52, 18 May 2024
physiologists study fundamental processes of plants, such as photosynthesis, respiration, plant nutrition, plant hormone functions, tropisms, nastic movements...30 KB (3,553 words) - 20:52, 18 May 2024 Botany (redirect from Plant biology)called plant science (or plant sciences), plant biology or phytology, is the science of plant life and a branch of biology. A botanist, plant scientist...138 KB (14,771 words) - 17:21, 26 October 2024
Botany (redirect from Plant biology)called plant science (or plant sciences), plant biology or phytology, is the science of plant life and a branch of biology. A botanist, plant scientist...138 KB (14,771 words) - 17:21, 26 October 2024- to activate chemical defenses against herbivores. It was the first plant hormone proven to be a peptide. Systemin induces the production of protein defense...15 KB (1,928 words) - 09:53, 3 December 2023
 "Effects of Salicylic Acid on the Bioproductivity of Plants". Salicylic Acid: A Plant Hormone. pp. 15–23. doi:10.1007/1-4020-5184-0_2. ISBN 978-1-4020-5183-8...31 KB (3,416 words) - 15:07, 8 October 2024
"Effects of Salicylic Acid on the Bioproductivity of Plants". Salicylic Acid: A Plant Hormone. pp. 15–23. doi:10.1007/1-4020-5184-0_2. ISBN 978-1-4020-5183-8...31 KB (3,416 words) - 15:07, 8 October 2024 Auxin (redirect from Rooting hormone)Native auxins Auxins (plural of auxin /ˈɔːksɪn/) are a class of plant hormones (or plant-growth regulators) with some morphogen-like characteristics. Auxins...45 KB (5,263 words) - 00:05, 12 September 2024
Auxin (redirect from Rooting hormone)Native auxins Auxins (plural of auxin /ˈɔːksɪn/) are a class of plant hormones (or plant-growth regulators) with some morphogen-like characteristics. Auxins...45 KB (5,263 words) - 00:05, 12 September 2024- Bioidentical hormone replacement therapy (BHRT), also known as bioidentical hormone therapy (BHT) or natural hormone therapy, is the use of hormones that are...74 KB (7,750 words) - 11:50, 3 July 2024
 Indole-3-butyric acid (category Plant growth regulators)decomposes before boiling. IBA is a plant hormone in the auxin family and is an ingredient in many commercial horticultural plant rooting products. Since IBA...7 KB (678 words) - 23:22, 13 April 2024
Indole-3-butyric acid (category Plant growth regulators)decomposes before boiling. IBA is a plant hormone in the auxin family and is an ingredient in many commercial horticultural plant rooting products. Since IBA...7 KB (678 words) - 23:22, 13 April 2024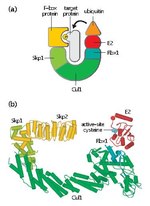 SCF complex (section Plant hormone signaling)cancer cells and cervical cancer cells to existing chemotherapies. The plant hormone auxin binds Tir1 (Transport Inhibitor Response 1). Tir1 is an Auxin...16 KB (2,021 words) - 19:31, 28 January 2023
SCF complex (section Plant hormone signaling)cancer cells and cervical cancer cells to existing chemotherapies. The plant hormone auxin binds Tir1 (Transport Inhibitor Response 1). Tir1 is an Auxin...16 KB (2,021 words) - 19:31, 28 January 2023- Alan M. Jones (section Plant Hormone Receptors)as a hormone receptor remains controversial among plant cell biologists because the structure and properties of ABP1 do not fit any animal hormone receptor...5 KB (695 words) - 12:09, 31 August 2024
 Cucurbita (redirect from Squash (plant))physiological, having to do with the age of the plant and whether it already has developing fruit. The plant hormones ethylene and auxin are key in fruit set...121 KB (10,840 words) - 00:42, 27 October 2024
Cucurbita (redirect from Squash (plant))physiological, having to do with the age of the plant and whether it already has developing fruit. The plant hormones ethylene and auxin are key in fruit set...121 KB (10,840 words) - 00:42, 27 October 2024 Gibberellic acid (category Plant hormones)or GA3) is a hormone found in plants and fungi. Its chemical formula is C19H22O6. When purified, it is a white to pale-yellow solid. Plants in their normal...7 KB (629 words) - 08:33, 19 July 2024
Gibberellic acid (category Plant hormones)or GA3) is a hormone found in plants and fungi. Its chemical formula is C19H22O6. When purified, it is a white to pale-yellow solid. Plants in their normal...7 KB (629 words) - 08:33, 19 July 2024 export. Programmed senescence seems to be heavily influenced by plant hormones. The hormones abscisic acid, ethylene, jasmonic acid and salicylic acid are...15 KB (2,074 words) - 17:39, 11 January 2024
export. Programmed senescence seems to be heavily influenced by plant hormones. The hormones abscisic acid, ethylene, jasmonic acid and salicylic acid are...15 KB (2,074 words) - 17:39, 11 January 2024 Deciduous (redirect from Deciduous plant)other. The cells are sensitive to a plant hormone called auxin that is produced by the leaf and other parts of the plant. When auxin coming from the leaf...18 KB (2,176 words) - 21:07, 12 September 2024
Deciduous (redirect from Deciduous plant)other. The cells are sensitive to a plant hormone called auxin that is produced by the leaf and other parts of the plant. When auxin coming from the leaf...18 KB (2,176 words) - 21:07, 12 September 2024- Gibberellin (category Plant hormones)Gibberellins (GAs) are plant hormones that regulate various developmental processes, including stem elongation, germination, dormancy, flowering, flower...37 KB (4,395 words) - 17:06, 26 October 2024
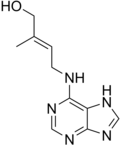 Cytokinin (category Plant hormones)Cytokinins (CK) are a class of plant hormones that promote cell division, or cytokinesis, in plant roots and shoots. They are involved primarily in cell...18 KB (1,978 words) - 22:50, 13 February 2024
Cytokinin (category Plant hormones)Cytokinins (CK) are a class of plant hormones that promote cell division, or cytokinesis, in plant roots and shoots. They are involved primarily in cell...18 KB (1,978 words) - 22:50, 13 February 2024
















- an article on: plant hormone Wikipedia plant hormone (plural plant hormones) (biochemistry) Any of many substances, produced by plants, that control or
- Hormonal contraception refers to birth control methods that act on the endocrine system. The original hormonal method—the combined oral contraceptive pill—was
- Volume 86 February 1915 (1915) Ductless Glands, Internal Secretions and Hormonic Equilibrium III by Fielding Hudson Garrison 1581066Popular Science Monthly
- period. Hormone supplements - Naturally the effects depend on the hormone but all in all these are beneficial but expensive. Plant growth hormone extracted