
Search results
There is a page named "Haplogroup L3 (mtDNA)" on Wikipedia
 Haplogroup L3 is a human mitochondrial DNA (mtDNA) haplogroup. The clade has played a pivotal role in the early dispersal of anatomically modern humans...40 KB (3,749 words) - 10:09, 11 July 2024
Haplogroup L3 is a human mitochondrial DNA (mtDNA) haplogroup. The clade has played a pivotal role in the early dispersal of anatomically modern humans...40 KB (3,749 words) - 10:09, 11 July 2024- Haplogroup H is a human mitochondrial DNA (mtDNA) haplogroup. The clade is believed to have originated in Southwest Asia, near present day Syria, around...51 KB (4,822 words) - 09:56, 11 July 2024
- descendant of the haplogroup L3. All mtDNA haplogroups found outside of Africa are descendants of either haplogroup N or its sibling haplogroup M. M and N are...47 KB (4,627 words) - 10:15, 11 July 2024
- In human mitochondrial genetics, Haplogroup C is a human mitochondrial DNA (mtDNA) haplogroup. Haplogroup C is believed to have arisen in East Asia some...59 KB (4,645 words) - 14:08, 26 June 2024
- Haplogroup J is a human mitochondrial DNA (mtDNA) haplogroup. The clade derives from the haplogroup JT, which also gave rise to haplogroup T. Within the...27 KB (2,697 words) - 11:35, 20 July 2024
- In human mitochondrial genetics, Haplogroup A is a human mitochondrial DNA (mtDNA) haplogroup. Haplogroup A is believed to have arisen in Asia some 30...63 KB (4,075 words) - 15:49, 18 July 2024
- Haplogroup K, formerly Haplogroup UK, is a human mitochondrial DNA (mtDNA) haplogroup. It is defined by the HVR1 mutations 16224C and 16311C. It is now...31 KB (3,034 words) - 10:06, 11 July 2024
 Haplogroup F is a human mitochondrial DNA (mtDNA) haplogroup. The clade is most common in East Asia and Southeast Asia. It has not been found among Native...49 KB (3,024 words) - 00:16, 19 June 2024
Haplogroup F is a human mitochondrial DNA (mtDNA) haplogroup. The clade is most common in East Asia and Southeast Asia. It has not been found among Native...49 KB (3,024 words) - 00:16, 19 June 2024- macro-haplogroup N, is a descendant of the haplogroup L3. All mtDNA haplogroups considered native outside of Africa are descendants of either haplogroup M...161 KB (14,828 words) - 00:48, 19 July 2024
 Haplogroup R is a widely distributed human mitochondrial DNA (mtDNA) haplogroup. Haplogroup R is associated with the peopling of Eurasia after about 70...45 KB (4,252 words) - 10:27, 11 July 2024
Haplogroup R is a widely distributed human mitochondrial DNA (mtDNA) haplogroup. Haplogroup R is associated with the peopling of Eurasia after about 70...45 KB (4,252 words) - 10:27, 11 July 2024 Z) Macro-haplogroup L is the most basal of human mtDNA haplogroups, from which all other haplogroups descend (specifically, from haplogroup L3). It is...23 KB (1,275 words) - 18:26, 26 June 2024
Z) Macro-haplogroup L is the most basal of human mtDNA haplogroups, from which all other haplogroups descend (specifically, from haplogroup L3). It is...23 KB (1,275 words) - 18:26, 26 June 2024 Haplogroup X is a human mitochondrial DNA (mtDNA) haplogroup. It is found in North America, Europe, Western Asia, North Africa, and the Horn of Africa...28 KB (3,032 words) - 15:44, 18 July 2024
Haplogroup X is a human mitochondrial DNA (mtDNA) haplogroup. It is found in North America, Europe, Western Asia, North Africa, and the Horn of Africa...28 KB (3,032 words) - 15:44, 18 July 2024- mitochondrial genetics, Haplogroup D is a human mitochondrial DNA (mtDNA) haplogroup. It is a descendant haplogroup of haplogroup M, thought to have arisen...110 KB (7,444 words) - 17:54, 26 April 2024
- Haplogroup T is a human mitochondrial DNA (mtDNA) haplogroup. It is believed to have originated around 25,100 years ago in the Near East.[citation needed]...59 KB (5,544 words) - 10:30, 11 July 2024
- human mitochondrial genetics, Haplogroup H5 is a human mitochondrial DNA (mtDNA) haplogroup descended from Haplogroup H (mtDNA). H5 is defined by T16304C...13 KB (1,411 words) - 14:12, 26 June 2024
- mitochondrial genetics, haplogroup E is a human mitochondrial DNA (mtDNA) haplogroup typical for the Malay Archipelago. It is a subgroup of haplogroup M9. Two contrasting...20 KB (1,434 words) - 14:12, 26 June 2024
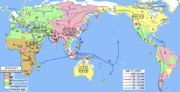 genetics, haplogroup Q is a human mitochondrial DNA (mtDNA) haplogroup typical for Oceania. It is a subgroup of haplogroup M29'Q. Haplogroup Q is a descendant...4 KB (438 words) - 14:11, 26 June 2024
genetics, haplogroup Q is a human mitochondrial DNA (mtDNA) haplogroup typical for Oceania. It is a subgroup of haplogroup M29'Q. Haplogroup Q is a descendant...4 KB (438 words) - 14:11, 26 June 2024- mitochondrial genetics, Haplogroup G is a human mitochondrial DNA (mtDNA) haplogroup. Haplogroup G is a descendant of haplogroup M. Haplogroup G is divided into...42 KB (2,318 words) - 14:09, 26 June 2024
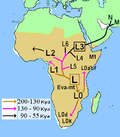 respectively. Haplogroups L5 (120 kya), L2 and L6 (90 kya), L4 (80 kya) and L3 (70 kya). The outgroup for mtDNA phylogeny of modern humans is the mtDNA of archaic...43 KB (4,268 words) - 12:09, 23 February 2024
respectively. Haplogroups L5 (120 kya), L2 and L6 (90 kya), L4 (80 kya) and L3 (70 kya). The outgroup for mtDNA phylogeny of modern humans is the mtDNA of archaic...43 KB (4,268 words) - 12:09, 23 February 2024 divisions for mtDNA haplogroups: African: L0, L1, L2, L3, L4, L5, L6 West Eurasian: H, T, U, V, X, K, I, J, W (all listed West Eurasian haplogroups are derived...36 KB (3,302 words) - 01:19, 3 July 2024
divisions for mtDNA haplogroups: African: L0, L1, L2, L3, L4, L5, L6 West Eurasian: H, T, U, V, X, K, I, J, W (all listed West Eurasian haplogroups are derived...36 KB (3,302 words) - 01:19, 3 July 2024








- lineages present in Africa only the female descendants of one lineage, mtDNA haplogroup L3, are found outside Africa. Had there been several migrations one