
Search results
There is a page named "Gas exchange" on Wikipedia
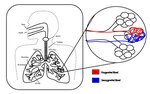 Gas exchange is the physical process by which gases move passively by diffusion across a surface. For example, this surface might be the air/water interface...57 KB (6,623 words) - 14:04, 30 September 2024
Gas exchange is the physical process by which gases move passively by diffusion across a surface. For example, this surface might be the air/water interface...57 KB (6,623 words) - 14:04, 30 September 2024- Cutaneous respiration (redirect from Cutaneous gas exchange)Cutaneous respiration, or cutaneous gas exchange (sometimes called skin breathing), is a form of respiration in which gas exchange occurs across the skin or outer...6 KB (608 words) - 14:50, 22 October 2024
- National Stock Exchange of India 5% – Oil and Natural Gas Corporation 5% – Adani Gas 5% – Torrent Gas 5% – GAIL Rest – Indian Energy Exchange and other Investors...7 KB (576 words) - 14:28, 24 September 2024
 Respiratory system (section Gas exchange)biological system consisting of specific organs and structures used for gas exchange in animals and plants. The anatomy and physiology that make this happen...95 KB (11,266 words) - 08:45, 28 September 2024
Respiratory system (section Gas exchange)biological system consisting of specific organs and structures used for gas exchange in animals and plants. The anatomy and physiology that make this happen...95 KB (11,266 words) - 08:45, 28 September 2024- gas-exchange cycles (DGC), also called discontinuous ventilation or discontinuous ventilatory cycles, follow one of several patterns of arthropod gas...16 KB (1,961 words) - 04:10, 19 June 2024
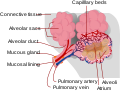 distensible cup-shaped cavities in the lungs where pulmonary gas exchange takes place. Oxygen is exchanged for carbon dioxide at the blood–air barrier between...27 KB (3,044 words) - 01:01, 21 June 2024
distensible cup-shaped cavities in the lungs where pulmonary gas exchange takes place. Oxygen is exchanged for carbon dioxide at the blood–air barrier between...27 KB (3,044 words) - 01:01, 21 June 2024 seamless market in natural gas used to fuel power stations. In the late 1990s, Sprecher acquired Continental Power Exchange, Inc. with the objective of...42 KB (4,326 words) - 20:58, 12 September 2024
seamless market in natural gas used to fuel power stations. In the late 1990s, Sprecher acquired Continental Power Exchange, Inc. with the objective of...42 KB (4,326 words) - 20:58, 12 September 2024 Extracorporeal membrane oxygenation (redirect from Extracorporeal gas exchange)heart and lungs are unable to provide an adequate amount of oxygen, gas exchange or blood supply (perfusion) to sustain life. The technology for ECMO...54 KB (5,673 words) - 20:57, 3 September 2024
Extracorporeal membrane oxygenation (redirect from Extracorporeal gas exchange)heart and lungs are unable to provide an adequate amount of oxygen, gas exchange or blood supply (perfusion) to sustain life. The technology for ECMO...54 KB (5,673 words) - 20:57, 3 September 2024- is the volume of air that is inhaled that does not take part in the gas exchange, because it either remains in the conducting airways or reaches alveoli...16 KB (2,227 words) - 12:58, 2 July 2024
- active movement that brings air into the lungs where the process of gas exchange takes place between the air in the alveoli and the blood in the pulmonary...7 KB (768 words) - 22:41, 21 June 2024
 Gas is one of the four fundamental states of matter. The others are solid, liquid, and plasma. A pure gas may be made up of individual atoms (e.g. a noble...52 KB (6,592 words) - 08:46, 7 September 2024
Gas is one of the four fundamental states of matter. The others are solid, liquid, and plasma. A pure gas may be made up of individual atoms (e.g. a noble...52 KB (6,592 words) - 08:46, 7 September 2024 walking along microtubules. A biological example of diffusion is the gas exchange that occurs during respiration within the human body. Upon inhalation...15 KB (1,776 words) - 08:15, 23 March 2024
walking along microtubules. A biological example of diffusion is the gas exchange that occurs during respiration within the human body. Upon inhalation...15 KB (1,776 words) - 08:15, 23 March 2024 fluorometer – gas exchange systems allow a more precise measure of photosynthetic response and mechanisms. While standard gas exchange photosynthesis...110 KB (11,776 words) - 21:08, 24 October 2024
fluorometer – gas exchange systems allow a more precise measure of photosynthetic response and mechanisms. While standard gas exchange photosynthesis...110 KB (11,776 words) - 21:08, 24 October 2024 for the direct exchange of gases between the internal tissues and atmosphere through the bark, which is otherwise impermeable to gases. The name lenticel...14 KB (1,627 words) - 13:38, 28 September 2024
for the direct exchange of gases between the internal tissues and atmosphere through the bark, which is otherwise impermeable to gases. The name lenticel...14 KB (1,627 words) - 13:38, 28 September 2024 Respiratory failure results from inadequate gas exchange by the respiratory system, meaning that the arterial oxygen, carbon dioxide, or both cannot be...18 KB (1,806 words) - 08:03, 29 September 2024
Respiratory failure results from inadequate gas exchange by the respiratory system, meaning that the arterial oxygen, carbon dioxide, or both cannot be...18 KB (1,806 words) - 08:03, 29 September 2024 of the GCMS results inconclusive. The gas exchange (GEX) experiment (PI: Vance Oyama, NASA Ames) looked for gases given off by an incubated soil sample...47 KB (5,320 words) - 21:21, 3 August 2024
of the GCMS results inconclusive. The gas exchange (GEX) experiment (PI: Vance Oyama, NASA Ames) looked for gases given off by an incubated soil sample...47 KB (5,320 words) - 21:21, 3 August 2024 Lung (section Gas exchange)and scorpions, which have book lungs). Their function is to conduct gas exchange by extracting oxygen from the air into the bloodstream via diffusion...106 KB (11,833 words) - 15:02, 22 October 2024
Lung (section Gas exchange)and scorpions, which have book lungs). Their function is to conduct gas exchange by extracting oxygen from the air into the bloodstream via diffusion...106 KB (11,833 words) - 15:02, 22 October 2024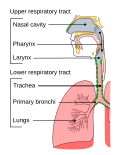 with the process of conducting air to the alveoli for the purposes of gas exchange in mammals. The respiratory tract is lined with respiratory epithelium...25 KB (3,061 words) - 17:22, 17 August 2024
with the process of conducting air to the alveoli for the purposes of gas exchange in mammals. The respiratory tract is lined with respiratory epithelium...25 KB (3,061 words) - 17:22, 17 August 2024- currency is exchanged for another Gas exchange, by diffusion across a surface Internet exchange point, allowing Internet service providers to exchange data Prisoner...2 KB (329 words) - 20:33, 30 June 2024
 epidermis of leaves, stems, and other organs, that controls the rate of gas exchange between the internal air spaces of the leaf and the atmosphere. The pore...37 KB (4,475 words) - 06:02, 22 October 2024
epidermis of leaves, stems, and other organs, that controls the rate of gas exchange between the internal air spaces of the leaf and the atmosphere. The pore...37 KB (4,475 words) - 06:02, 22 October 2024













- A Real Solution for High Gas Prices by Barack Obama/Podcasts 49023A Real Solution for High Gas PricesBarack Obama/Podcasts Hello, this is Senator Barack
- is the process that moves air in and out of the lungs to facilitate gas exchange with the internal environment, mostly to flush out carbon dioxide and
- is the exchange of gases (oxygen and carbon dioxide) between inhaled air and the blood. INTERNAL RESPIRATION, which is the exchange of gases between